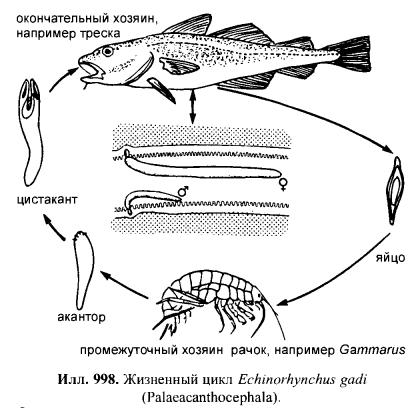
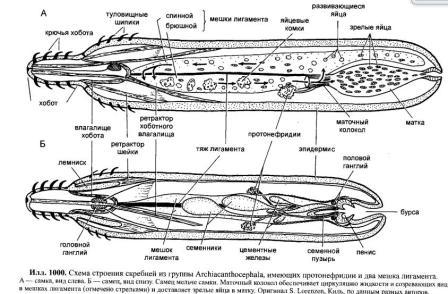
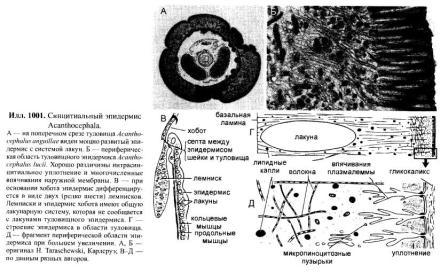
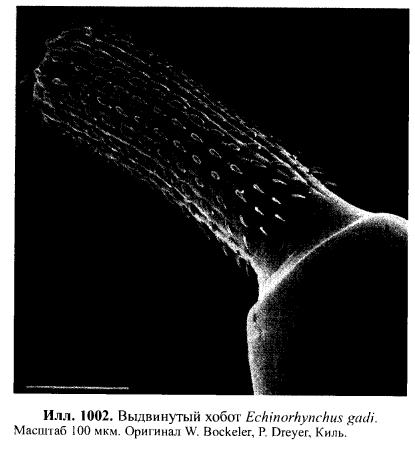
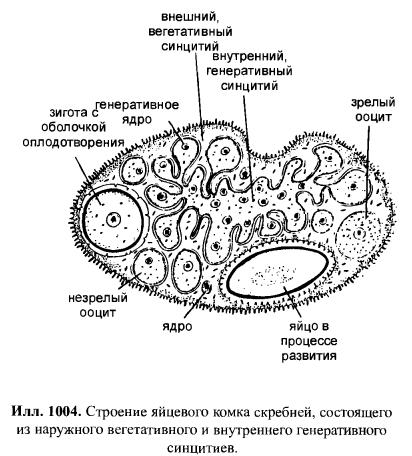
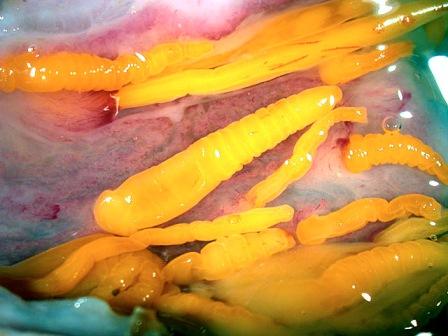
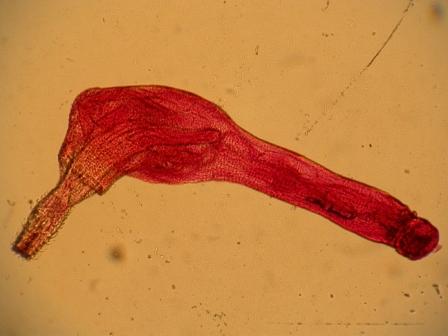
Acanthocephala, Скребни Все скребни являются кишечными паразитами с обязательной сменой хозяев. Их окончательными хозяевами служат водные или наземные позвоночные, а промежуточными хозяевами — ракообразные и насекомые. Часто присутствует резервуарный хозяин, в котором развитие паразита не происходит. Смена хозяина не сопровождается сменой поколений паразита. Группа насчитывает примерно 1 100 видов. Размеры скребней варьируют от 2 мм до 70 см и у большинства видов не превышают нескольких сантиметров; самки всегда крупнее самцов. Мелкие виды паразитируют в основном в рыбах, а более крупные — в птицах или млекопитающих. В последнее время представители Acanthocephala стали использоваться как биоиндикаторы для определения содержания свинца в воде: скребни, паразитирующие в рыбах, накапливают свинец в тканях гораздо интенсивнее, чем их хозяева. Строение. Тело подразделено на хобот, шейку и туловище. Хобот способен втягиваться в тело и выдвигаться вперед; он гомологичен хоботку коловраток из группы Bdelloida, но не гомологичен интроверту других представителей Nemathelminthes. Хобот несет направленные назад крючья, служащие для закрепления в стенке кишечника хозяина (отсюда происходит латинское название группы — буквально «колючеголовые»). Крючья формируются из базальной ламины эпидермиса. Вытянутое туловище у многих видов Palae- и Eoacanthocephala покрыто шипиками, которые — в отличие от крючьев хобота — представляют собой склеротизированные выросты эпидермиса. В редких случаях, например у Mediorhynchus taeniatus, туловище имеет поверхностную (ложную) сегментацию. Кишечник отсутствует на всех стадиях жизненного цикла, и питание паразита осуществляется внекишечно (через стенку тела). Число и расположение соматических клеток абсолютно постоянно (эвтелия), в то время как половые клетки размножаются в течение всей жизни скребня. Большинство тканей имеют синцитиальную природу. Подвижные реснички есть только в протонефридиях (которые имеются у представителей одного семейства — Oligacanthorhynehidae); кроме того, сперматозоиды скребней снабжены жгутиками. Характерной чертой скребней является наличие в теле нескольких разнокачественных полостей. Кроме собственно полости тела, разделенной на две части, в примитивном состоянии имеются два мешка лигамента, а также обширная система эпидермальных лакун. Определение брюшной и спинной сторон тела, не составляющее проблемы для большинства представителей Bilateria, в случае скребней является предметом дискуссии. У этих животных отсутствуют ротовое и анальное отверстия, а гонады открываются терминально; таким образом, отсутствуют важнейшие внешние признаки, маркирующие дорсовентральную полярность. Было установлено, что две стороны тела иногда различаются по степени развития шипиков, а при изгибании тела одна из сторон обычно становится вогнутой, другая — выпуклой. Оказалось также, что две стороны тела хорошо различаются по топографии внутренних органов, но какую из них следует считать брюшной, а какую — спинной, долгое время оставалось неясным. Ответ на этот вопрос, однако, может быть найден в рамках гипотезы о близком родстве скребней с коловратками из группы Bdelloida. У последних гонады всегда располагаются вентрально, а головной мозг всегда лежит дорсально от втянутого хоботка. Такое расположение органов характерно для всех Nemathelminthes, и в дальнейшем изложении мы будем предполагать, что оно свойственно и Acanthocephala. Эпидермис представляет собой синцитиальный эпителий. У молодых особей этот синцитий содержит 6-20 ядер, число и расположение которых видоспецифично. В течение жизни эти ядра становятся высокополиплоидными; у мелких видов они приобретают лопастную форму и достигают 2 мм в диаметре, а у многих крупных видов распадаются на фрагменты (так наз. амитоз). Снаружи эпидермис покрыт слоем гликокаликса толщиной около 1 мкм, который обеспечивает избирательный обмен веществ с окружающей средой. Периферическая зона эпидермиса уплотнена подобно тому, что наблюдается у коловраток, но не образует жёстких пластин. Это интрасинцитиальное уплотнение пронизано многочисленными каналами, которые представляют собой впячивания наружной мембраны эпидермиса. Похожие впячивания образует также и базальная ламина эпидермиса. Кроме этого, для эпидермиса скребней характерно развитие собственной интрасинцитиальной системы лакун, состоящей из продольных и кольцевых главных каналов и многочисленных анастомозирующих дополнительных каналов. Впячивания наружной мембраны служат для поглощения питательных веществ. Лакунарная система передней части тела (хобота и шейки) отделена от системы туловища. Эпидермис содержит многочисленные волокна, которые в периферическом слое располагаются, преимущественно, параллельно к поверхности тела и образуют войлокоподобное сплетение, а в толще эпидермиса формируют радиальные связки, лежащие между обширными каналами лакунарной системы. В области шейки эпидермис образует два (редко шесть) мешковидных выпячивания (лемниска), которые вдаются назад во внутреннюю полость туловища. Лемниски окружены мышцами-ретракторами и играют важную роль в работе гидравлической системы хобота, а также в его закреплении в стенке кишки хозяина. Под эпидермисом располагается мускулатура, состоящая из наружного слоя кольцевых мышц и внутреннего слоя продольных мышц. Оба слоя довольно тонкие и образованы трубчатыми синцитиальными мышцами. В стенке влагалища хобота проходят перекрещивающиеся мышечные волокна. Их антагонисты находятся вне стенок влагалища: это шейные ретракторы, прикрепляющиеся изнутри к кожно-мускульному мешку, и ретракторы хоботного влагалища и собственно хобота, которые также отходят от кожно-мускульного мешка. Работа мышц-антагонистов возможна благодаря гидроскелетной функции полости тела. При сокращении одной из групп мышц давление в полости тела увеличивается и растягивает те части тела, которые оказывают меньшее сопротивление давлению. Органы чувств развиты крайне слабо, что связано с эндопаразитическим образом жизни скребней. Отдельные сенсиллы располагаются на вершине хобота, по бокам шейки и в области полового отверстия. Обширная полость тела ограничена снаружи мышцами кожно-мускульного мешка, а изнутри — стенками мешков лигамента, и представляет собой псевдоцель. На уровне влагалища хобота эта полость разделяется на «целом хобота» и «туловищный целом». Значительную часть последнего занимают спинной и брюшной мешки лигамента. Тяж лигамента, лежащий в горизонтальной плоскости, разделяет эти мешки на значительном протяжении (но не полностью). Мешки лигамента тянутся от влагалища хобота до выводного протока гонад. У Palaeacanthocephala стенки мешков значительно редуцированы, так что их полости объединяются между собой и с полостью тела. Тяж лигамента можно рассматривать как рудимент кишки. По расположению и строению он соответствует рудименту кишки самцов коловраток из группы Monogononta. Церебральный ганглий лежит дорсально во влагалище хобота. Он состоит из очень небольшого числа нервных клеток (у большинства видов 73-86). От ганглия отходят два главных латеральных нервных ствола, которые тянутся в туловище. У самцов в районе пениса имеется пара ганглиев примерно из 30 нейронов; у самок соответствующие ганглии отсутствуют. Органы выделения отсутствуют у скребней, паразитирующих в рыбах или других водных позвоночных. Среди других форм протонефридии имеются только у Oligacanthorhynchidae, паразитирующих в наземных позвоночных. Протонефридии расположены двумя группами в дистальной части половой системы и открываются в клоаку вместе с половыми протоками. Скребни раздельнополы. Гонады у обоих полов располагаются в брюшном мешке лигамента, прикрепляясь к его дорсальной стенке. У самцов имеются два семенника, выводные протоки которых вместе с так называемыми цементными железами открываются на вершине пениса внутри половой бурсы. Женская половая система скребней имеет уникальное строение. Один или два яичника еще у личинки распадаются на яйцевые комки, которые затем свободно плавают в брюшном мешке лигамента (у Archi- и Eoacanthocephala) или в полости тела (у Palaeacanthocephala). Каждый яйцевой комок состоит из двух синцитиев. Наружный вегетативный синцитий несёт на поверхности многочисленные микроворсинки и получает питательные вещества из окружающей жидкости; во внутреннем генеративном синцитии находятся диплоидные ядра будущих ооцитов. Развитие ооцитов начинается с того, что генеративные ядра вместе с окружающими участками цитоплазмы погружаются в толщу наружного синцития и созревают, получая из него желток и другие вещества. Затем яйца отшнуровываются от внутреннего синцития и остаются в толще наружного синцитиального слоя. Размножение и развитие При копуляции самец охватывает задний конец тела самки выпяченной бурсой, вводит пенис во влагалище и выделяет сперматозоиды, которые затем активно мигрируют к яйцевым комкам. Копуляция может происходить неоднократно. После внедрения сперматозоида в ооцит про-исходят мейоз и кариогамия. Зигота имеет округлую форму; в ней сразу же начинается эмбриональное развитие, в котором обнаруживаются черты спирального дробления. Проследить судьбу отдельных бластомеров в ходе развития невозможно, поскольку бластомеры довольно рано сливаются в синцитий. Развитие яйца протекает в теле самки до полного созревания личинки; к этому времени яйцо приобретает веретеновидную форму (у Palaeacanthocepha1а), а его оболочка становится многослойной и твёрдой. Незадолго до окончания развития яйцо отделяется от яйцевого комка. Уникальным для группы является механизм сортировки яиц с помощью особого маточного колокола. Колокол лежит в брюшном мешке лигамента у входного отверстия матки. Ритмично сокращаясь, он всасывает жидкость вместе с плавающими в ней яйцами и нагнетает её в сортировочную камеру. Последняя сообщается с маткой щелью определённого размера, пройти через которую могут только узкие, веретеновидные зрелые яйца. Незрелые яйца округлой формы не проходят через щель, а выводятся в спинной мешок лигамента, движутся по нему вперёд вместе с током жидкости, а затем попадают обратно в брюшной мешок и снова подносятся к маточному колоколу. Важная функция колокола состоит в поддержании циркуляции жидкости в мешках лигамента, что обеспечивает перенос веществ между различными частями тела. У Palaeacanthocephala стенки мешков лигамента значительно редуцированы и циркуляция распространяется на весь туловищный отдел полости тела: Из матки яйца выводятся в просвет кишки хозяина и вместе с его экскрементами попадают во внешнюю среду. Яйца скребней весьма устойчивы к неблагоприятным условиям. В соответствии с паразитическим образом жизни продукция яиц у скребней очень высока. Например, в теле самки гигантского скребня Macracanthorhynchus hirudinaceus (длиной до 70 см) содержатся миллионы яиц, из которых ежедневно откладывается около 80 тысяч. У некоторых видов Ео- и Palaeacanthocephala оболочка яйца содержит сахара, которые служат приманкой для промежуточных хозяев. В кишке промежуточного хозяина (всегда Arthropoda) из яйца выходит личинка — акантор, которая активно проникает в стенку кишки и далее, через несколько недель, в полость тела хозяина. Гемоциты хозяина реагируют на личинку как на инородное тело, образуя вокруг неё тонкую капсулу. Внутри неё акантор продолжает развитие и превращается в акантеллу, уже имеющую втяжной хоботок. Полностью развитая акантелла обозначается как цистакант. Эта личинка продолжает питаться за счёт промежуточного хозяина, но не развивается до тех пор, пока промежуточный хозяин не будет съеден окончательным. Тогда цистакант выходит из капсулы и прикрепляется хоботом к стенке кишки окончательного хозяина, превращаясь во взрослую особь. При попадании в организм резервуарного хозяина цистакант также выходит из капсулы, но затем пробуравливает стенку кишки хозяина и снова инкапсулируется. При среднем уровне заражения скребни обычно не вызывают значительных повреждений. Человек заражается скребнями очень редко; теоретически заражение гигантским скребнем может произойти при употреблении в пищу личинок майского жука. Систематика Монофилия Acanthocephala хорошо подтверждается следующими уникальными признаками: (1) эпидермис с обширной системой лакун; (2) способный выпячиваться хобот с крючьями, которые развиваются из базальной пластины эпидермиса; (3) наличие мешков лигамента; (4) наличие маточного колокола у самок. Эвтелия соматических тканей Acanthocephala позволяет предположить, что их пред¬ки были микроскопически мелкими животными и обладали строгим постоянством клеточного состава. Это предположение подтверждается также тем, что у крупных видов скребней ядра эпидермиса не делятся митотически, а распадаются на фрагменты. Так как по всем данным паразиты происходят от свободноживущих предков, а не наоборот, и что микроскопически мелкие предки скребней тоже были свободноживущими. Аналогичная картина наблюдается у нематод, где крупные зоопаразитические виды происходят от мелких паразитических форм, а последние, в свою очередь — от микроскопически мелких свободноживущих предков. Ближайшими родственниками скребней считают коловраток из группы Bdelloida. В пользу этого можно привести следующие аргументы: (1) эпидермис Acanthocephala и Rotatoria имеет уникальное интрасинцитиальное уплотнение и пронизан каналами — впячиваниями апикальной мембраны; (2) лемниски присутствуют только у скребней и коловраток (прежде всего, Bdelloida); (3) и скребни, и Bdelloida имеют хобот, выдвижение которого происходит сходным образом; этот хобот отличается от интроверта Priapulida, Loricifera и Kinorhyneha тем, что не несет ротового отверстия; (4) ретракторы хобота Acanthocephala и Bdelloida имеют сходные ультраструктурные особенности; (5) по строению и положению в теле тяж лигамента скребней соответствует рудименту кишки самцов коловраток из группы Monogononta. Эти данные заставляют отвергнуть широко распространенное мнение о близком родстве скребней и приапулид, в основе которого лежит ошибочная интерпретация хобота скребней как интроверта. Что же касается филогении внутри Acanthocephala, то каждый из трёх таксонов скребней в своё время рассматривали как наиболее примитивный (это отражено в их названиях: греч. palaios = старый; eos = заря [начало]; archaios = древний). На основании родства с коловратками можно заключить, что в пределах Acanthocephala мелкие формы, паразитирующие в водных животных, должны быть примитивнее, чем крупные скребни, обитающие в наземных хозяевах. Эта точка зрения отражена в классификации, приведённой ниже. 1. Eoacanthocephala Мелкие виды. Окончательными хозяевами служат почти исключительно рыбы. Самки имеют два мешка лигамента, дорсальный и вентральный, с полностью развитыми стенками. Протонефридии отсутствуют. Neoechinorhynchns rutili, самки длиной 5-10 мм, в кишечнике пресноводных рыб. Промежуточные хозяева-— рачки из группы Ostracoda и личинки вислокрылок рода Sialis. 2. Palaeacanthocephala Мелкие или средней величины скребни. Окончательные хозяева — рыбы, амфибии, водоплавающие птицы и другие позвоночные, связанные с водной средой. Главные стволы лакунарной системы эпидермиса лежат латерально. У самок имеется только один мешок лигамента, возникающий в результате объединения спинного и брюшного мешков. Стенки мешков неполные, поэтому яйца свободно плавают в полости тела. Протонефридии отсутствуют. Echinorhynchus gadi, самки длиной 4-8 см, часто встречается в кишечнике трески; промежуточными хозяевами служат рачки Amphipoda. — Е. truttae, самки длиной 2 см, частый паразит форели; промежуточные хозяева — рачки Gammarus (Amphipoda). — Асаnthocephalus anguillae, самка длиной 1-3,5 см, обитают в кишечнике различных пресноводных рыб, промежуточный хозяин — водяной ослик Asellus aquaticus. 3. Archiacanthocephala Средней величины или крупные скребни. Окончательными хозяевами служат наземные позвоночные. Главные стволы лакунарной системы эпидермиса проходят дорсально и вентрально. Самки имеют два мешка лигамента, дорсальным и вентральным, с нормально развитыми стенками. Протонефридии имеются только в семействе Oligacanthorhynchidae, виды которого отличаются особенно крупными размерами. Macracanthorhynchus hirudinaceus, гигантский скребень, длина самки до 70 см, в кишечнике свиней; промежуточными хозяевами являются личинки жуков из группы Lamellicornia, например майского и июньского хрущей, а также бронзовок.
Источники Зоология беспозвоночных. Под ред. В. Вестхайде и Р. Ригера. Т. 1-2. М., 2008